

和谐:从基因说
2024-06-18 10:13:02
来源:西部决策网
分享到



生命是基因的天使,又是基因的奴仆。负荷生命的基因之河永久的流淌,记载着过去,昭示着未来。人类圈创建的基因选择机制,削弱了自然选择,正在重构人与自然关系。
——是为题记

生命是基因之河,基因是生命天书。地球上所有的生命皆以基因为密码。人类已是地球霸主,但也只是生命长河中的又一匆匆过客。基因是生命与自然关系的载体,深深地永久的刻印着曾经的过往。面向未来,构建人与自然和谐共生关系,要从阅读生命天书、解读基因密码开始。
一、基因及其复杂度
经历三年“新冠疫情”,人们对“新冠病毒”“做核酸”刻骨铭心。“新冠疫情”的日子已经过去,从与病毒抗争到彼此适应,我们的生命永远留下了“新冠病毒”侵入和改造的痕迹。这一留痕,是否会世代相传,尚未可知。不过,从既往病毒侵入史中,科学家已经发现,病毒参与了生物进化。具有逆转录功能的病毒,可以把基因整合进宿主基因组,所有多细胞生物基因组都有病毒插入的基因。在人类基因组中,有8-10%的DNA为病毒所贡献。人类不断建立更加完善的值得信赖的免疫系统和应答机制。
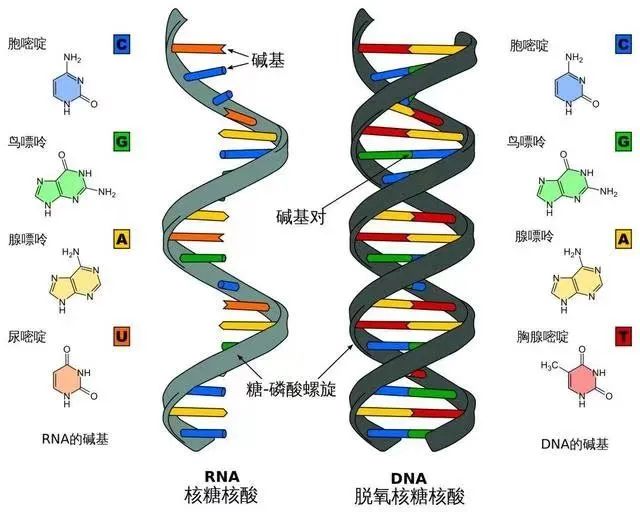
根据分子结构特征,将“核酸”分为脱氧核糖核酸(DNA)和核糖核酸(RNA)。根据携带“核酸”类型,亦将病毒分为DNA病毒和RNA病毒。新冠病毒是RNA病毒,由一个核酸长链和蛋白质外壳构成。病毒具有部分生命特征却算不上完整的生命,因而被定义为非细胞生命形态。因没有进行新陈代谢所必需的生命系统,不能自我复制,病毒必须借助特定的宿主细胞完成复制、转录和转译等生命活动。三年疫情反复“做核酸”,就是从宿主细胞采样,检测来自病毒的核糖核酸(RNA),以阳与阴表示有与无。
生物学家从细胞学视野(暂无能力依基因分类),将已知生命划分为三个域:细菌、古菌和真核生物。真核生物即复杂的多细胞生物,包括动物、植物和真菌。真核生物的遗传物质集中在细胞核(细胞的“中控室”)中,且有提供能量的细胞器——线粒体或是捕获光子能的叶绿体,包含内质网在内的细胞膜网络,以及由肌动蛋白和微管蛋白组成的细丝构成的细胞骨架。细菌、古菌属于原核生物,其细胞结构和功能要简单许多,不仅没有细胞核,包裹DNA的蛋白,也缺少线粒体(叶绿体)及胞质结构等。
生物进化过程就是生物复杂度上升过程。40亿年前(亦说38亿年前),海洋热泉生化反应创建了生命密码,生成了细菌、古菌。20亿年前,细菌、古菌融合共生(细菌套细菌)产生了真核细胞,形成了早期的真核生物。也就是说,真核生物是原核生物——细菌、古菌细胞融合发展而来的,先是形成了结构功能完整的单细胞生物,再形成了复杂的多细胞生物。这是一次基因复杂度大提升,也是一次生物大革命。2015年,科学家对深海环境进行研究时,发现了有着真核生物基因的奇怪古菌。2017年,科学家在另外几种古菌中发现了类似的基因。这被认为是生命系统发生树中最接近真核生物的一类古菌。
DNA是所有生命的共有密码。DNA是一连串的合成RNA和蛋白质所需的遗传密码。DNA和RNA有序管控着细胞内同时进行着的各种生化反应。在生命构建与运行过程中,DNA永久存续,而RNA只是DNA的阶段表达,当使命完成后即行销毁。几乎所有的生物使用同样的遗传密码,病毒也不例外。遗传密码从原始生命生化系统协同演化而来。遗传密码的形成是生命诞生的重要标志。这是生物共祖论的遗传密码证据。
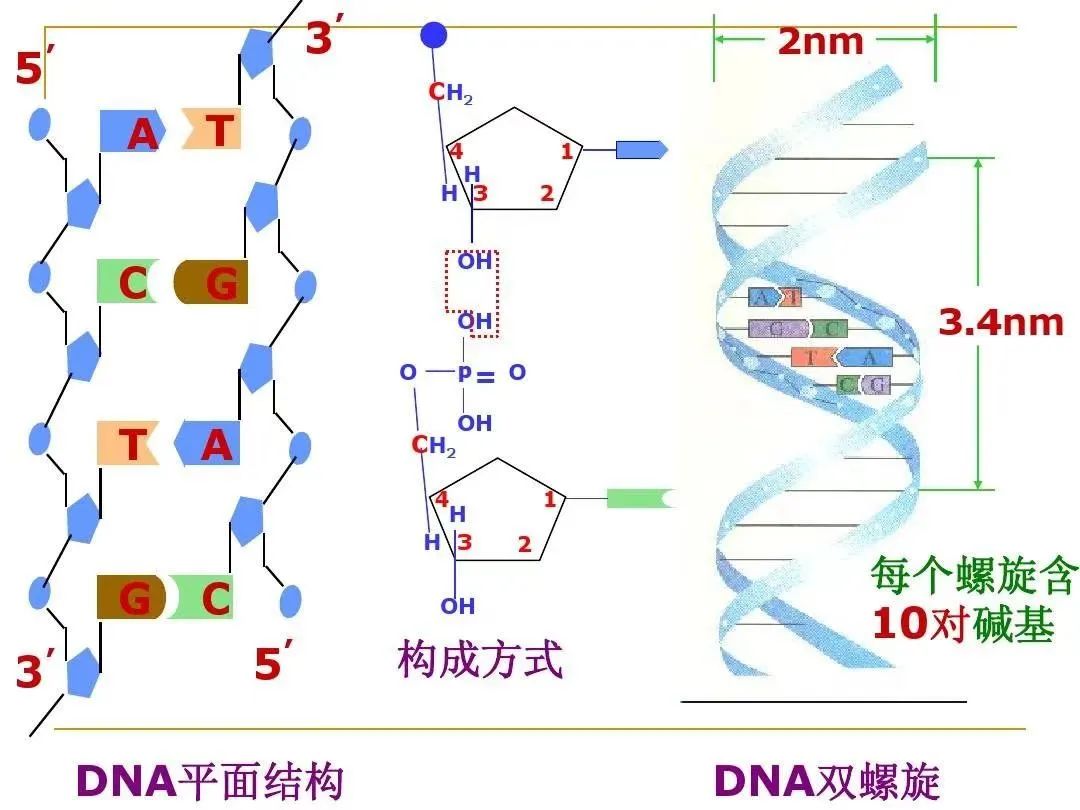
生物复杂度集中在DNA上。原核生物——细菌、古菌的遗传物质DNA通常为裸露的双螺旋长链。真核细胞的遗传物质DNA上升为蛋白质包裹的双螺旋长链,且在细胞核中以染色体为载体。一条染色体即是一条DNA载体,亦是一个DNA包裹。包裹DNA的材料是蛋白质。真核细胞在进行有丝分裂或减数分裂时,细胞核中的染色体可见。在不分裂时,染色体不可见。原核细胞DNA裸露无包裹,也就没有可见的染色体。
精诚合作是基因组基本原则。原核生物不仅细胞结构简单,世代较短,基因组较小,一般只含有数百至数千个基因。真核生物细胞结构复杂,世代变长,基因组变大,一般拥有数万个基因。真核生物细胞核内染色体上的基因是基因主体,线粒体、叶绿体内也有少量基因,核内核外基因合在一起即是基因组。基因一体,和合共生,不是单打一、自顾自,自私内耗。基因组、一家人,密切配合、通力协作。精诚合作是基因组经过漫长岁月磨砺进化而来的基本原则。生物复杂度的底层逻辑是基因组复杂度。基因组复杂度是生物复杂度的重要标尺。
有性生殖是提升生物复杂度的灵巧机制。生命之初,皆是底底道道的自我复制——无性生殖,世代差异微小。有性生殖是真核生物的一大发明,也是自然选择的有利突变。性构筑起复杂生命的第一支柱,也提供了发展的第一动力。基因组精妙细微,深藏不漏,肉眼不可观。然而,基因组用尽心思,塑造了可直观的性之外形,铺平了两性细胞融合发展的“性选择之路”,提供了基因复杂度升级、生命向复杂化进化的灵巧机制。自有性生殖以来,加速了世代差异,推动了生物进化,地球上的生命变得丰富多彩、复杂多样。人类已拥有高度复杂的基因组,且得有性生殖之真缔,成为性高度复杂的生物。是不是已经抵达复杂度峰值,有待深入观察。
基因只是DNA的片段。基因是DNA长链上具有遗传能力的一个片段,全部片段总合在一起即是基因组。一条DNA长链上包含多个基因,每一个基因只是其中一个遗传密码单元。DNA的一个片段就是一个密码单元,也就是一个基因。这一片段一单元一基因,就是能够产生一条多肽链或RNA所需的核苷酸序列。一个基因的大小,也是一个密码单元的大小,它取决于所产生的一条多肽链或RNA所需的核苷酸序列的大小。一个基因约为200-2000个遗传密码子。
遗传密码子就是3联碱基符。3个连续的核苷酸组合在一起形成1个遗传密码子,常用3个碱基符号表示,比如ATG。“核苷酸”单位中的“核”指碱基,“苷”指五碳糖,“酸”指磷酸。碱基是遗传密码的核心。碱基总共5种,分别是腺嘌呤(A)、鸟嘌呤(G)、胞嘧啶(C)、胸腺嘧啶(T)和尿嘧啶(U)。A 、G、C和T存在于DNA中,A、G、C和U存在于RNA中。碱基通过共价键与核糖或脱氧核糖的1位碳原子相连而形成核苷。核苷再与磷酸结合形成核苷酸。核苷酸共有C、H、O、N、P5种化学元素。千姿百态的生命形式,其遗传物质都是5种碱基、5种化学元素。
1号染色体是最霸气的生物大分子。核甘酸聚合而成的核酸分子通常很大,DNA分子是已知的最大的单个生物分子,RNA分子则是较小的生物分子(最小的RNA分子仅21个核苷酸)。人体细胞有23对(46条)染色体,其中22对常染色体,1对性染色体。减数分裂时产生的配子染色体数目由46条减半为23条,碱基对由62亿减半为31亿。人体1号染色体是最霸气的生物大分子,包含2.46亿个碱基对,3141个基因,约占人类DNA的8%。破译1号染色体,将为研究治疗癌症、帕金森氏症和老年痴呆症等350余种疾病提供科学指引。2号染色体仅次于1号染色体,2.37亿个碱基对,1888个基因,包含重要的发育调控基因簇。科学研究认为,人类2号染色体似是黑猩猩两对染色体拼合而成(染色体突变),这使人类染色体数目(23对)比黑猩猩染色体数目(24对)少了1对,也成为人类与黑猩猩有共同祖先的基因证据。
基因是小纳米级的存在。想一想细胞的大小、精子的大小,再想一想细胞核的大小,染色体在细胞核中,DNA在染色体上,62亿碱基对、6万余基因,以及精子卵子配对,受精卵开始新的生命历程,无限精妙皆在其中。不要说用肉眼了,一般的显微镜也无法观察到DNA,更别说只是其中一个片段的基因了。1989年,美国科学家用“扫描隧道显微镜”直接观察到了脱氧核糖核酸的双螺旋结构和尺寸。DNA两条多核苷酸链反向平行盘绕所生成的双螺旋直径2.0nm,相邻碱基对之间的轴向距离0.34nm,每个螺旋的轴距3.4nm。
RNA才是演化的起点?现代生命的“中心法则”:DNA—RNA—蛋白质是一个相互依存、无法撕裂的圆圈,人们难以甄别谁才是演化的起点。而RNA世界学说认为,地球的大多数生物以DNA作为遗传分子,同时用RNA辅助完成一些功能。DNA出现之前,可能存在一个“RNA世界”。这一理论推测,在生命起源的某个时期,生命体仅由一种高分子化合物RNA组成,兼具DNA和蛋白质功能。遗传信息的传递建立于RNA的复制,其复制机理与当今DNA复制机理相似,作为生物催化剂的、由基因编码的蛋白质还不存在。在上亿年进化过程中,RNA逐渐将其携带遗传信息的功能传给了DNA,将其功能分子的功能则传给了蛋白质。

复杂基因操控复杂的纳米机器人。蛋白质是生命中的纳米机器人,与生命的产生、存在和消亡密切相关。蛋白质质量决定生命质量,没有蛋白质就没有生命。蛋白质的基本单位是20种氨基酸,而20种氨基酸千变万化的组合,则听命于遗传信息。生物复杂程度上升,意味着所需要的蛋白质种类增加,而蛋白质由基因控制的纳米机器人。生物复杂度上升,必然要求纳米机器人增加,要求基因编码及非编码序列随之增加,要求负责指导和调控转录的DNA区段――调控序列的长度随着增长。真核生物中编码蛋白质的基因通常不连续,内含子增加致使编码一个基因所需的DNA长度增加。同时,基因间隔区是基因组上与RNA和蛋白质表达无关的那部分DNA,如假基因、转座元件等,基因间隔区序列数目增加导致基因密度降低。这在一定程度上进一步加速提升了基因的复杂度。
基因复杂度原本只是意外事件的累积。因为各种各样的原因,在DNA复制的过程中会出现一定数量的差错(如同文本誊写错误),这样的意外事件被称之为基因突变。基因的每一次复制,都会产生一定数量的突变。随着基因组规模增大,基因复制精度有所提高,但复制错误在所难免,基因突变是复杂生物必须面对的生命常态。基因突变是完全随机事件,而自然选择则是优胜劣汰。基因突变分为三种,有害突变、中性突变和有利突变。一般而言,自然选择的有性生殖会严格把控基因的质量。(1)中性突变影响轻微,长期存留在种群中并无多大妨碍。可以说,有性生殖为中性突变设置了黄灯区。(2)有性生殖为有害突变设置了道道红灯。携带有害突变基因的个体,呈现生存缺陷,有的精卵不能融合,有的妊娠早期流产,即使出生也多会夭亡,抑或在婚配市场遭弃,不会留下后代,有害基因随之清除。有害突变难以越过有性生殖自然选择这一关。如果有害突变率过高,将会带来“突变灾难”,危及物种群体安全。(3)有性生殖对有利突变一路绿灯。携带有利突变基因的个体获得了生存优势,可以留下更多后代而逐步发展壮大,直至携带有利基因的个体成为种群主体。长期而缓慢的有利突变累积,推动基因复杂化、生物复杂化。这看似漫无目的的随机的突变,经过自然选择机制,将自然环境变迁的信息,也整理记录在基因组内。这是一部基因差错版摇身一变成为基因改良版的史诗,它铸就了生物进化的基本原理。基因复制出现差错只是个体的偶然事件,却是群体的必然事件。大自然向各物种公平提供生态机会,自然选择主导的生物进化永远在路上。

二、生态位及其竞争性在流淌不息的基因之河中,在环环相扣的生命链条上,基因竞争是永恒法则。所有生命都在接受基因竞争的挑战。基因总在拼命复制自己,复制中有差错有成败,客观上形成了竞争局面。生物进化没有幕后设计师,只有未来引导师。自然选择,说到底就是利用基因竞争机制,引导基因发展方向。自然为生命发展提供了生态位——生态资源、生态机会。然而,自然提供的生态资源、生态机会具有多样性稀缺性,需要竞争获得。自然选择就是自然引导。引导基因竞争具有多样性稀缺性的生态资源、生态机会,继而形成多样性的基因组、多样性的物种和多样性的生态系统。
40亿年来,地球上出现过的所有生命统合在一起,构成了一部完整的地球生命史,其主题归结为一句话便是:从无主之地到众生乐土。科学家估计,地球上曾经出现过的全部生物,其中99%已经成为生物先躯。这1%的现存生物是同时代地球生态圈的共生物种,与99%的先躯生物有着千丝万缕的联系。99%的先躯生物已经淌过了基因之河,把面向未来的有利基因和全部的生态位留给了1%的现存生物。1%的现存生物并不是孤立的存在,而是45亿年地球环境变迁、生物突变、自然选择、生命接续的演化结果。曾经出现的每一种生物,都曾利用过生态机会,使用过生态资源,都曾在生态位上占有过一席之地。钩沉99%生物的历史积淀,必将为1%生物的未来前景提供历史镜鉴。
在地球形成不久,地球环境中就形成了构建生命的基材——有机化学物。由此,打开了生命的机遇之门。大约40亿年前,出现了后来地球生命的共同祖先——细菌、古菌。这一人类肉眼观察不到的微小生命体,不仅能够一代接着一代永续复制自身的结构,还能重现自身的功能。奥妙在于世代相传的DNA,永久保存着其结构与功能的密码。迄今为止,人们依然不能确切地知道原核生物究竟是怎么形成的。看似最简单的生命,其复杂程度也超过了当今的科学技术极限,人们穷尽一切,始终无法再现其形成过程。原核生物早在40亿年前就已经形成的史实,广为人知、无可争议。那个时候,地球尚是无主之地。在20亿年的漫长岁月里,整个地球——从海洋到陆地都在迎接有机世界来临,等待着成为细菌、古菌的占领区。而原核生物的行动非常谨慎,一直把行为规范在海洋里、水境中。大约20亿年前,发生了一次破天荒的微生物革命。在海洋的某个场景中,古菌吞噬了细菌,但并没有将其消化,而是合二为一、一体共生。这被称为内共生现象,其有力证据是,真核细胞(共生细胞)的细胞核中的DNA像是来自古菌,真核细胞的线粒体中残留的DNA像是来自细菌。由此,原核细胞融合发展形成了真核细胞,地球生命史迈上了真核生物发展的新阶段。动物、植物和真菌(本意是蘑菇)随之而来,生生不息。
真核细胞在海洋中默默演化的10亿年里,并没有留下向多细胞生物发展的地质证据。成冰纪(8.5亿年-6.3亿年前),地球被冻成雪球,加速了矿物入海,客观上增加了海洋营养物质。埃迪卡拉纪(6.35亿年前 ~ 5.41亿年前),动物已开始增长,主要是刺胞动物,包括水母类和海鳃类,以及三叶动物、盾形动物、多孔动物、环节动物、节肢动物、栉水母动物等。然而,进入寒武纪时(距今5.42亿年—4.85亿年前),海洋中生命大爆发。这一时期的地层中充斥着各种各样的海洋生物。当今存在的几乎所有大型动物种类,不同的身体结构和组织结构,早在寒武纪时就已经形成,其中也包括脊椎动物。及此,可以说生物已经完美的占领了海洋。在海洋由无主之地发展成为众生乐园之时,地球陆地依然是无主之地,提供了广阔的生态机会。

陆地生态系统构建过程,就是生物竞争陆地生态位过程。植物是陆地生态系统的主体,也是登陆的先行生物、基础生物。与动物一样,植物也经历了从简单到复杂、从水生到陆生的进化过程。大约40亿年前,光合细菌是植物的祖先。大约在20亿年前,开始出现多种藻类,如绿藻、红藻、棕藻等。当初的光合细菌并不是绿色的,但绿色光合细菌最终胜出,是因为它能够更有效地利用阳光而顺利通过了自然选择。光合作用的一大副产品是排出了氧气,因光合作用开启了地球大氧化过程,同时也为动物的出现创造了条件。原始的单细胞绿藻在原始海洋中进化为多细胞藻类,这是植物进化的起点。奥陶纪(4.8-4.4亿年),低等海生植物继续发展,淡水植物可能已经出现,逐渐积累起开拓陆地生态位的力量。志留纪(4.4-4.1亿年)晚期,绿色植物成功登陆,初始的高等植物——裸蕨植物首次出场。由绿藻进化出苔藓,由苔藓进化出蕨类,陆地植物接二连三。从泥盆纪(鱼类的时代4.1-3.6亿年)开始,陆地生态事件就成为地球生态事件的主旋律。泥盆纪早期,裸蕨类植物兴盛繁荣。中期后,腕足类和珊瑚发育、原始菊石、昆虫出现。晚期原始两栖类、迷齿类出现,蕨类植物和原始裸子植物出现。大约持续1亿年植物登陆历程,绿色占领了地球陆地。到了石炭纪(3.6-2.8亿年),陆地上形成了大规模的森林,给煤的形成创造了有利条件。生活在陆上的昆虫,如蟑螂类和蜻蜓类,是石炭纪突然崛起的一类陆生动物。二叠纪(2.99-2.5亿年)是古生代最后一纪,由蕨类进化出裸子植物(松柏类),银杏、苏铁亦开始发展。裸子植物用种子繁殖,具有种子和木质化的主干,对干旱和寒冷环境具有较强适应能力。白垩纪(1.44-0.66亿年),由裸子植物进化出被子植物(开花植物),并成为今天植物界的主角。被子植物通过花粉、雄蕊和雌蕊等生殖器官进行繁殖,具有更高的成活率和种类多样性。至此,植物已经进化出复杂的细胞结构,如叶绿体、木质部、造枝组织等,表现出种类繁多的叶、茎、根、花和果实,光合作用、呼吸作用、传递养分、抗病和防御机制日趋完善。与植物相比,动物占领陆地生态位过程波澜壮阔,但在时间上要相对晚一些。大约3.5亿年前,海洋动物经由湿地抵达陆地,从水生到两栖再到陆生,逐步占领陆地。再后来,出现细菌、真菌等营腐生生活的微生物,它们将动植物的尸体残肢分解还原为二氧化碳,水和无机盐,回归无机环境中,形成了无机物—植物—动物—微生物—无机物的生态环境大循环。至此,陆地提供的生态机会被广泛利用,陆地也由无主之地变为众生乐土。
从海洋到陆地,从无主之地到众生乐园、众生乐土,意味着地球生命领地扩张过程已经结束,生态位竞争进入了全新阶段。生态位竞争是基因竞争的时间空间表达。我们可以把生命在海洋扩张、以占据海洋空缺生态位为主的时代,称之为1.0时代的生态位竞争,把生命在陆地扩张、以占据陆地空缺生态位为主体的时代,称之为2.0时代的生态位竞争,在海洋和陆地占据空缺生态位过程结束后,进入以生态位分化为主的新阶段,即是3.0时代的生态位竞争。
3.0时代的生态位具有特定的内涵。(1)空间位。每个物种都有其特定的栖息地和生活范围,这就是它们的生态空间。比如,不同鸟类生活在森林不同层次,有的在树冠层,有的在中间层,有的则在地面上,形成生态空间差异。(2)时间位。生态位会随季节变化,包括海拔高度、经纬度的迁徙移动。(3)营养位。亦可称能量位。在食物链、能量链上的位置以及相互作用。狮子处于生态顶端,以动物为食,羚羊以植物为食,植物以无机物为食,植物—羚羊—狮子各有其营养位。(4)共生位。当生态资源稀缺时,共生物种进行竞争;当生态资源充足时,促进生态位分化,建立新的生态位共生关系。生态位多样性是生物多样性的基础。
同理,生物竞争就是生态位竞争。通过各自占据不同的生态位避免大范围的冲突对抗,以维持生态系统的稳定性、多样性和持续性。生态位竞争循环四大原则:(1)适配原则。寻求与自身生理需求适配的生态位是生物本能。(2)排斥原则。已经取得适配生态位的生物是既得利益者,极力排斥既得利益竞争者。(3)开拓原则。自然界总有生态空隙、无主之地,等待新的开拓者和占领者,增加生态位多样。(4)精致原则。与生命复杂度提升一样,生态系统亦由简单向复杂演进,生态位竞争势必走向精致化。

生态位竞争是维持生物多样性和生态系统稳定性的重要机制。通过生态位竞争,物种能够有效地利用生态资源和生态机会,不断提升生物种群和生态系统生产力。一般而言,生物竞争表现为种间竞争和种内竞争。种间竞争的本质就是生态位竞争。种内竞争不属于生态位竞争,但会向生态位竞争传导压力,并影响生态位竞争力。
人类是生物,高度参与了生态位竞争。生物竞争既有种间竞争,又有种内竞争。本文不关注人类种内竞争(制度之争、科技之争、产业之争),只关注人类与其他物种间的生态位竞争。自从进入猎业时代以来,人类就取得了无可比拟的竞争优势。无数物种纷纷向人类让出了世代相传的生态位。或者换种积极的说法,人类充分发挥竞争优势,开拓和占领了越来越多的生态位。人类圈日益扩大,越来越多的生态位转化为人类生态位。同时,也意味着被人类挤出生态位的生物越来越多,失去生态位的生物越来越多,无家可归、走投无路、求生无门的生物越来越多。无数曾经繁盛的物种,因被挤出生态位而沉没于基因之河。无可争议,经历猎业时代、农业时代、工业时代,人类在与其生物的竞争中已经获得了全面胜利。人类先后成为陆地霸主、海洋霸主、空中霸主,如今已全面称霸陆海空。在40亿年地球生命史上,曾经遇见过不少霸主,想一想不可一世、称霸陆地长达1.5亿年的恐龙。但是,与人类在一万年时间内就全面称霸陆海空的霸业成就相比,恐龙之霸业显然逊色太多、不足道哉!
在3.0时代的生态位竞争中,人类获得大胜并左右了生态位竞争大趋势。目前,人类以胜利者姿态规划未来生态位,从生态位竞争者一跃而成为生态位供给者、创建者,从而开启了生态位竞争的4.0时代。人类圈制定并实施国土空间规划,城镇空间、农业空间原本是生态位中的“白菜心”,也是人类早已抓在手心心的生态位,自然要守牢用好,决不能拱手让出。自然生态空间原本是生态位中的“边角料”,因占用成本过高,人类曾视之为荒野,将其继续留给自然,供其他物种使用,亦合情合理,纳入规划序列则号称合规。纵然如此,这也是破天荒的大事件。在人类圈力量盛隆,生物圈人类化的滚滚洪流中,其他物种仅存的生态位能够纳入国土空间规划,完全得益于人与自然和谐共生之心。人类圈通过制定实施国土空间规划,稳定生态位秩序,稳定生物多样性,稳定人与自然关系,这也是值得众生颂扬的人类生态位美德。

三、从自然选择到天人双选
人,因思想而伟大。人类最独特的能力莫过于思想。当今,流行“人与自然和谐共生”的思想。不少人立马会联想到“天人合一”的思想。的确,二者一脉相承,逻辑相通。然而,“天人合一”是结草为庐、自然环境中产生的思想,“人与自然和谐共生”则是高楼林立、人造环境中的思想。在自然环境中谈人与自然关系,与在人造环境中谈人与自然关系,当然具有不同意境。人类不可能放弃智能家室,退居茅草居所。今时之“人与自然和谐共生”,决非回归旧时的“天人合一”,而是构建新时代生态文明新形态。
人从自然来,自然生成了生态生产力。生态生产力是原生生产力,社会生产力是次生生产力。人类利用生态生产力,从生态系统中采集渔猎,获得最初的发展动力。人类的起源具有随机性、偶然性,但人类文明发展于生态生产力较高,且能便利获得采集产品、渔猎产品的森林、湿地生态位上。人类利用生态生产力过程中,不断向生态系统施加了压力。人类喜欢占据粗大的象牙,加速了生长粗大象牙的大象退却,生长细小象牙的大象成为象群的主导者。人类喜欢占据了高大通直的木材,加速了森林中高大通直树木退却,低矮弯曲的树木成为森林的主角。凡是人类喜欢占据的动植物,大致都会承受巨大的压力,面临退出生态位的风险,而那些不被人类青睐的动植物,反而获得了更多的生态位。
人类不仅在利用自然,还在改造自然。如此这般,令自然遭受双重损失。驯化是人类主导下的进化,既是利用自然又是改造自然。人类喜食小麦、水稻、玉米、谷子、土豆以及梨、枣、桃、杏、梅、柑橘、苹果、核桃……就将其驯化栽培,视其为农作植物,清除森林、草原植物,开辟耕地,经营农作。人类喜欢狗、猪、鸡、牛、羊、鱼、兔、马、驴……亦将其驯化饲养,视其为家养动物,建立各种各样的养殖场、放牧场。1万年间,农作植物、家养动物发生的变化,显然不是自然选择,而是人类选择。与农作植物、家养动物相对应,湿地、森林、草原中的原生动植物则被视之为野生动植物。前者是关系家庭生计的,已经驯化并被占有的,而后者则是无关家计的,属于自然而然的。有人认为,前者与人类已经结成了生计伙伴,也即是与人类友好的动植物。友好的动植物支持人类生计,人类帮助友好的动植物扩张领地范围。在人类的助力下,农作植物、家养动物得到更多的生态位(也算是人类生态位)。没错,这形成了极为融洽的双向互馈。然而,有得必有失。人类大规模占领生态位,阻断了生态位分化的进化之路。受害的是农地上的土著——野生动植物,它们失去了世袭罔替的生态位,也失去了进化的光明前景。工业化以来,人类利用自然和改造自然的强度增加,人类生态位持续扩大,加速了野生动植物生态位流失。目前,世界各地的昆虫数量在以惊人的速度下降,预计未来几十年,40%的昆虫物种将会灭绝。昆虫数量减少,或是因为农业集约化,抑或是农药的使用,又或是昆虫生态位丧失?有资料显示,人类生产和消费食物的方式已经造成了80%以上的生物多样性丧失。
人类生态位扩张,野生动植物生态位流失,带来了极为深刻而广泛的生态学问题。人类新得生态位,即是野生动植物新失生态位。人类新占据生态位上的原有动植物,因丧失生态位而沉没坠落基因之河。早在猎业时代,人类就干掉了大型猎物。受人类生态位胁迫,所有生物呈现小型化趋势。再联想到地理大发现后的全球生物大交换,人类圈全球化深度影响了生态位分化和生物进化过程。长期以来,生物遗传学专注于DNA复制、基因突变以及自然选择机制(简称“天选”)。现在的问题是,自从人类获得生态位超级竞争力以来,纵然DNA复制、基因突变的规则并变化,但自然选择机制已大为弱化。人类的选择已经十分重要,深刻的影响并削弱了自然选择的力量。1万年来,人类按照自己的想法,一直用各种技术(包括太空育种)选择培育农作植物、家养动物品种,用各种方法治疗基因突变带来的疾病,卓有成效的建立起基因突变应对机制和基因选择机制(简称“人选”)。人类圈不仅向生物圈施加了直接的压力,人类活动还引起全球气候变化、生态环境变化,间接的影响到自然选择。比如,碳排放引发的全球气候变暖,可能使适应暖化气候的基因突变有更多机会通过自然选择。但是,如果人类长期脱离体力劳动,且仅习惯居住在几乎恒温的居室里,将不利于耐暖化气候的基因突变,与自然选择走上相反的道路。生态环境污染使得部分物种永久丢失,当生态环境修复后,那些丢失的物种却再也回不来了。

这里,我们不妨把自然选择看作“市场机制”,是生物进化的“无形推手”,人类选择则如同“计划机制”,构成生物进化的“有形推手”。“天选”与“人选”的关系,就如同“市场”与“计划”的关系。人类圈广泛影响到生物进化机制,“天选”与“人选”两种机制都在发挥作用。在自然生态系统中,自然选择一以贯之。而在人类圈,人类选择已占主导。面向未来,基因选择的天平上,“天选”与“人选”孰强孰弱?尚不宜断言。
人类不仅按照自身意志选择其他物种,也做出一系列制度安排,驯化和选择人类自己。制度的前体是文化。文化亦是进化推手。婚配选择就是基因选择,婚姻制度就是基因制度。在人类制度史上,婚配制度始终居于重要地位。人类婚配竞争压力远远超过了其他物种,不仅有与其他物种一样的生物竞争,还包含着独特的文化竞争、社会竞争。不仅有外貌长相的自然选择,还有金钱财富、文化素养、社会地位的选择。郎才女貌、门当户对是古今中外、流传千年的婚配市场基本规则。进入信息时代,婚配市场数据增加,而婚配选项也在增加,反而导致婚姻选择困难症。不少人的生殖力,在观望、徘徊、等待中渐渐流失。
日益复杂的产权制度、交易制度、教育制度、就业制度、社保制度,对婚配市场和生殖力也具有重要影响。人类内部生态位(社会生态位)竞争已成进入智业时代,经济结构越来越复杂,科技门槛越来越高,新质生产力层出不穷,社会生态位持续分化,极限挑战人类的认知能力、创新能力。应对社会生态位分化与竞争的通用策略,就是把更多的资源分配在学习知识、掌握技能、增强本领和专注事业上,而不是用在生育后代上。社会选择了“好学”之人。“好学”替代“好斗”的社会进步机制,也会引起基因选择响应。生有涯而知无涯,以有涯随无涯,殆已!于无形之中,人类生殖力集体流失,极为宝贵的有利突变也随之灭失。

人类成为生物进化的重要推手,这也是地球霸主地位的重要象征。21世纪已经进入基因组时代,正在迎接多组学或生命组学时代的到来。基因科技已经在加速产业化,未来农业、精准医学、生物制造构成基因产业化发展的三大关键赛道。基因科技+人工智能,基因选择机制加速“突变”,必将重构生命进化秩序。人类以天赋异禀搏击基因之河!兹事体大!关系万千生命的未来,也关系人类的未来。
人类经历猎业时代、农业时代、工业时代,正在进入智业时代。人类的生态位堪称“巨无霸”!地球上已经没有任何物种可以挑战人类的霸主地位。人类的力量显得过于庞大,以致于生物世界已经背离了达尔文多样性越来丰富的进化之路。我们的世界变得更加单调了而不是更加多样了,更加枯燥乏味了而不是更加生动有趣了!我们的学历越来越高,知识越来越多,本领越来越强,寿命越来越长,这一切究竟为什么?人生而自由,然而无往不在枷锁之中。我们比祖辈得到的更多,但我们比祖辈更幸福吗?未来的人类,更像生物一般挥洒自如,还是更像机器一样有条有理?
人类不是生物进化的终点,也不是生物进化的终结者。当人类淌过基因之河,让出生态机会、生态资源时,者大自然将会重构地球生态系统,再度演绎更为灿烂的生命故事。大概与恐龙想不到后恐龙时代的世界一样,人类也想不到人类基因之河干涸之后的世界。以我们已经获得的智慧而言,地球上已经没有人类生态位竞争者。人类大规模退回生态位的唯一可能的原因,就是人类生殖力流失,人口数量呈现减少趋势。人类圈崩解之后,地球霸主让位于新人类!无论如何,人类都将成为未来生命的作者、创造者。但愿人类是一位卓越的作者、伟大的创造者!(文/党双忍)

作者注:本文为《和谐》系列10。和谐是全人类共有价值观。部分插图源自网络。2024年6月18日成作于磨香斋。

返回顶部